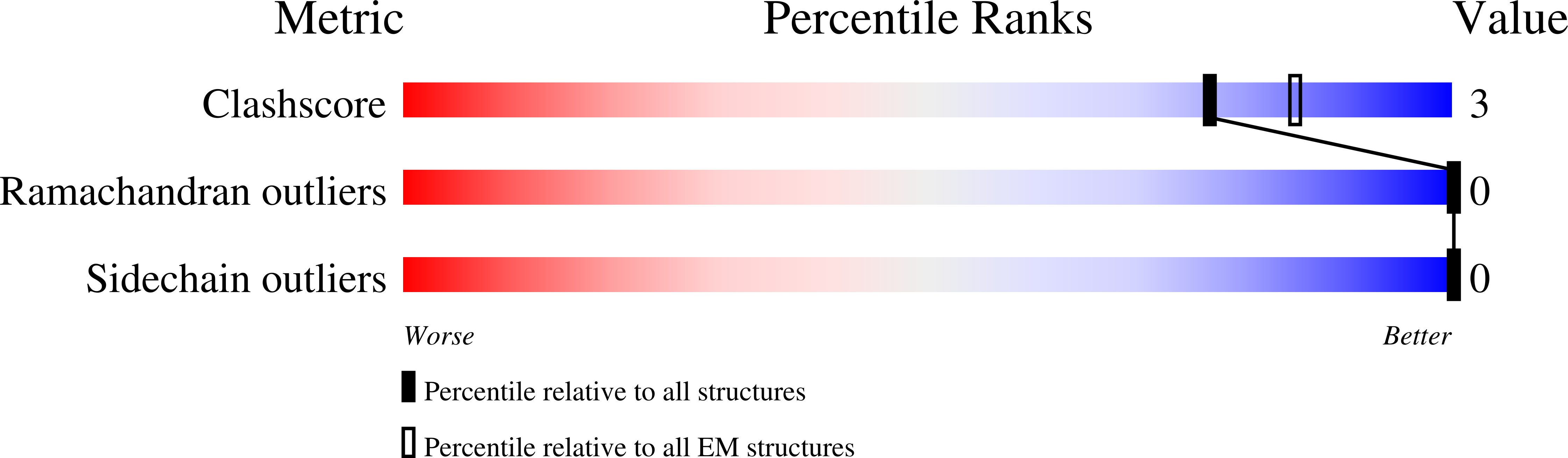
Deposition Date
2024-07-23
Release Date
2025-04-23
Last Version Date
2025-05-21
Entry Detail
PDB ID:
9CSD
Keywords:
Title:
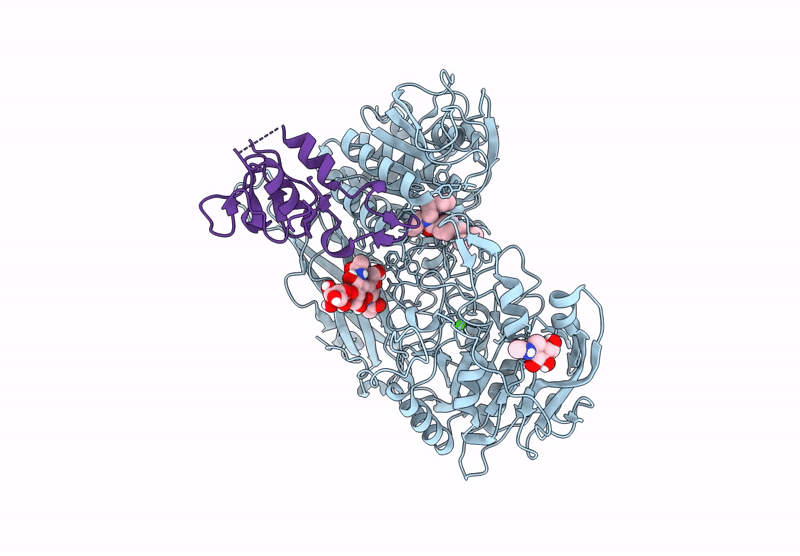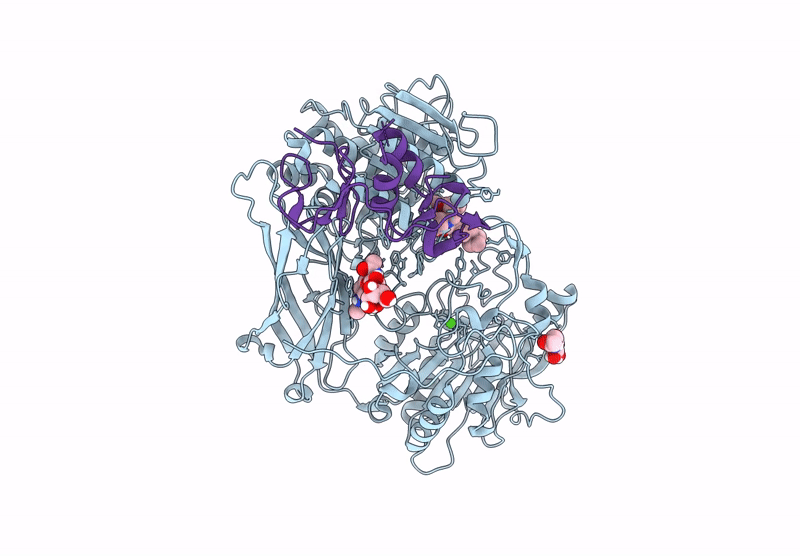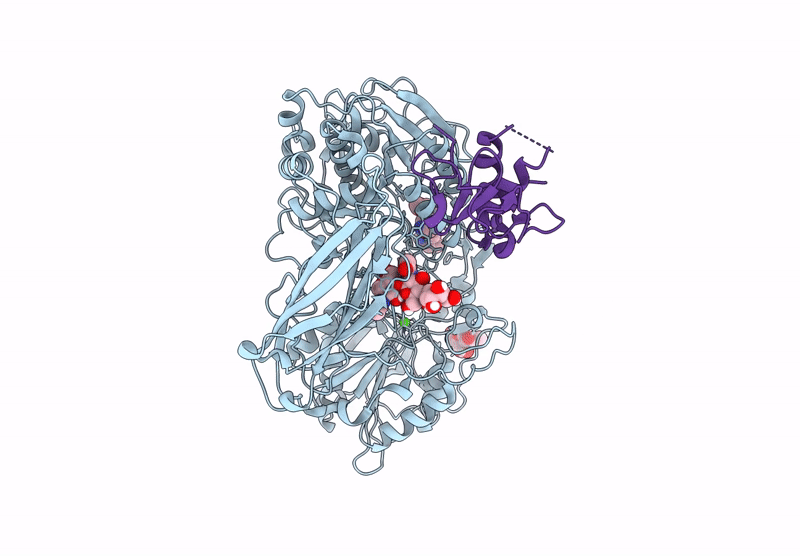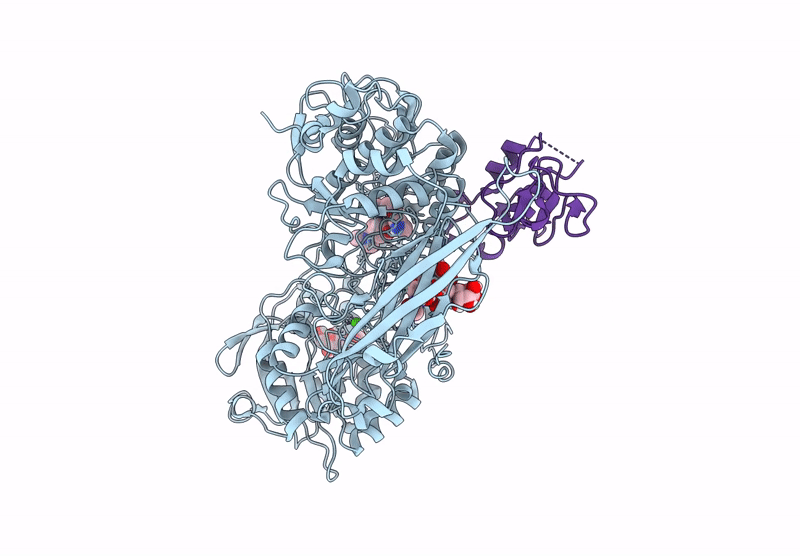
The Ectodomains of SPRING and S1P with the inhibitor PF-429242
Biological Source:
Source Organism(s):
Homo sapiens (Taxon ID: 9606)
Expression System(s):
Method Details:
Experimental Method:
Resolution:
2.40 Å
Aggregation State:
PARTICLE
Reconstruction Method:
SINGLE PARTICLE