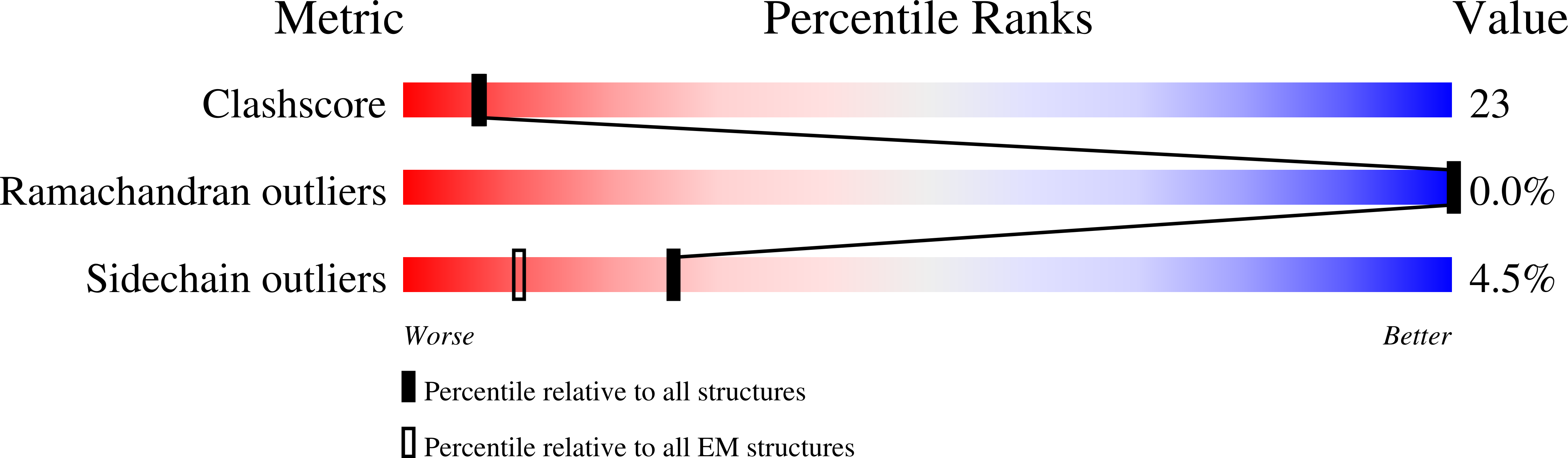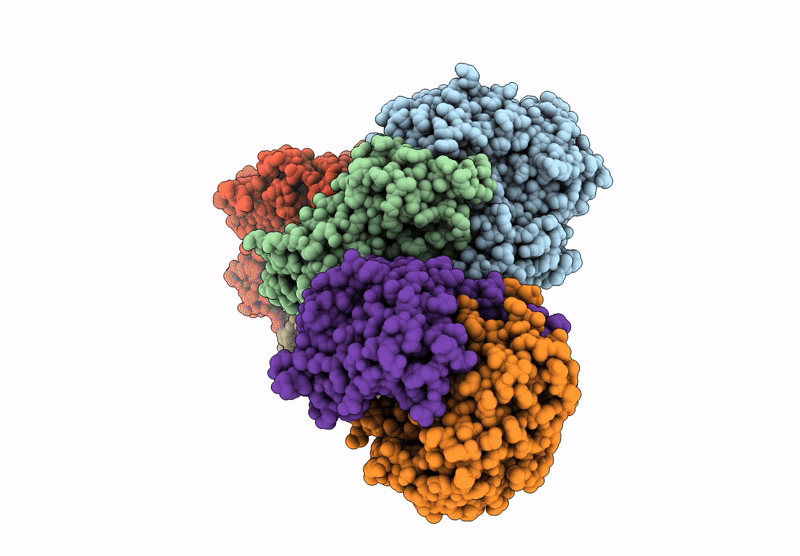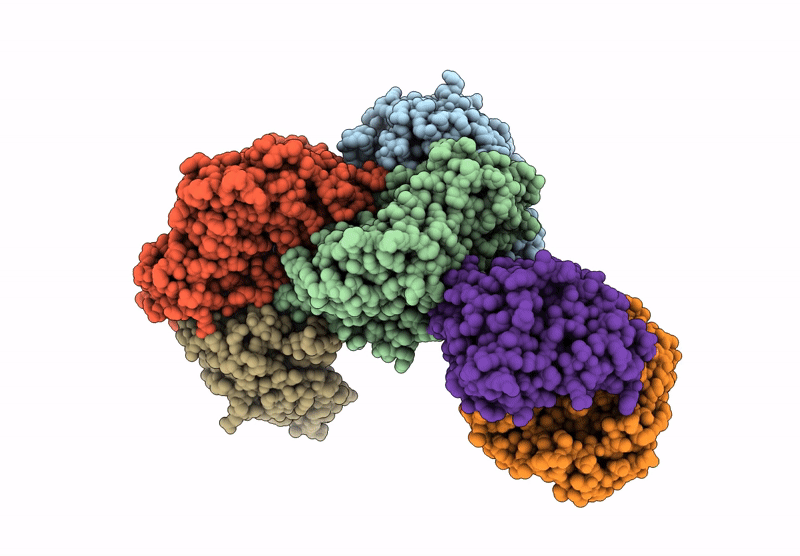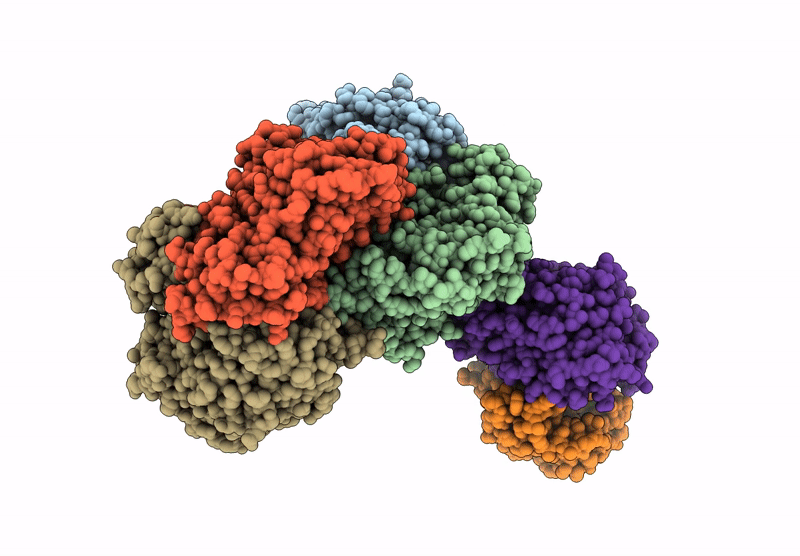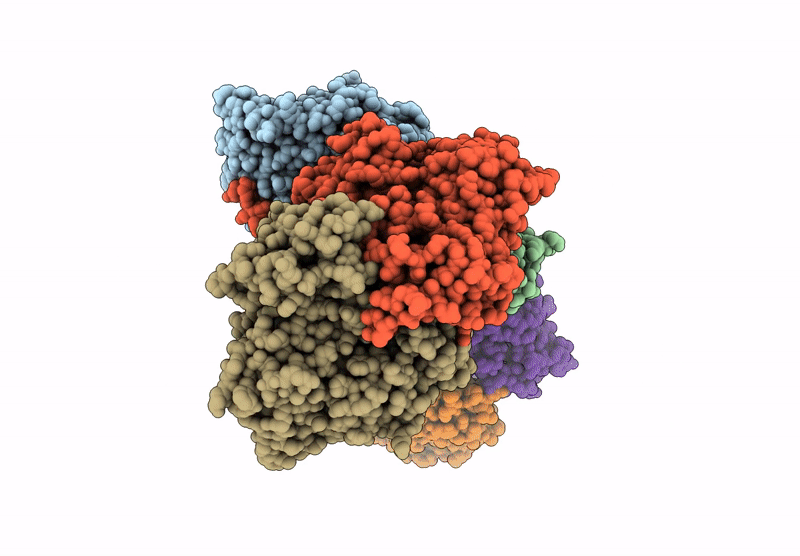
Deposition Date
2024-01-18
Release Date
2025-04-30
Last Version Date
2025-06-04
Entry Detail
Biological Source:
Source Organism(s):
Cereibacter sphaeroides (Taxon ID: 1063)
Expression System(s):
Method Details:
Experimental Method:
Resolution:
3.82 Å
Aggregation State:
PARTICLE
Reconstruction Method:
SINGLE PARTICLE