
Deposition Date
2023-12-04
Release Date
2024-12-11
Last Version Date
2025-06-25
Entry Detail
PDB ID:
8RBF
Keywords:
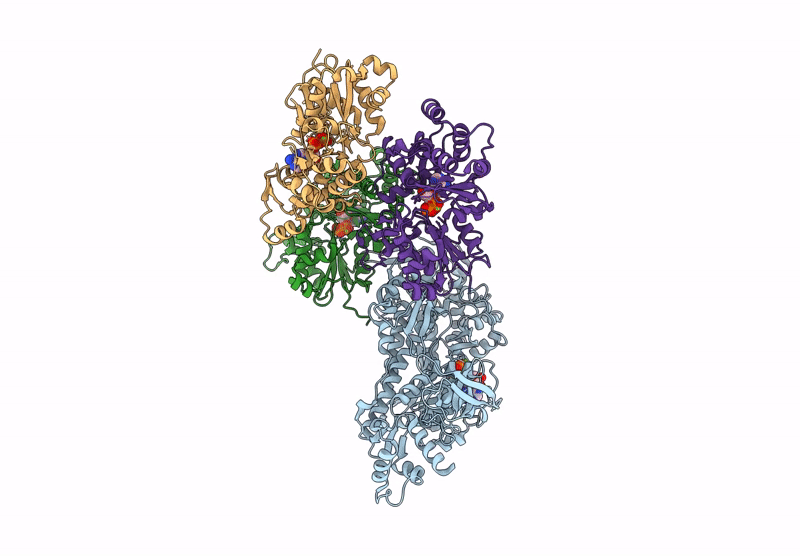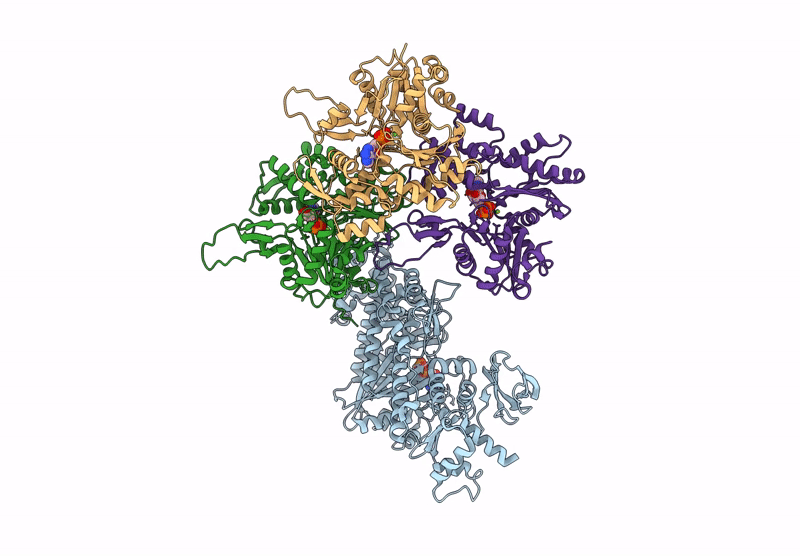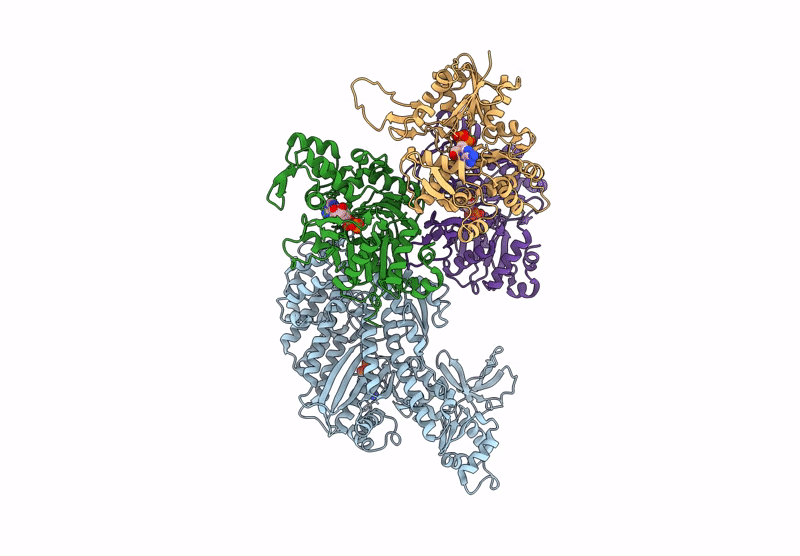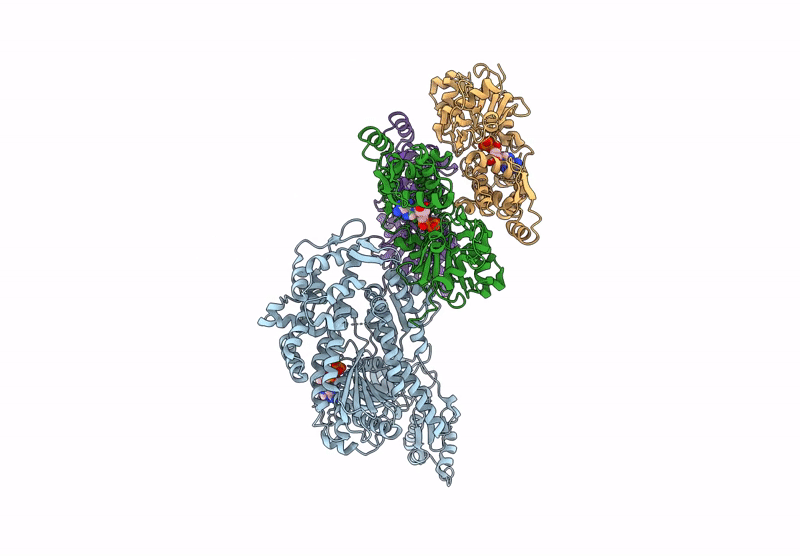
Title:
CryoEM structure of the post-powerstroke actomyosin-5a complex
Biological Source:
Source Organism(s):
Mus musculus (Taxon ID: 10090)
Oryctolagus cuniculus (Taxon ID: 9986)
Oryctolagus cuniculus (Taxon ID: 9986)
Expression System(s):
Method Details:
Experimental Method:
Resolution:
4.20 Å
Aggregation State:
FILAMENT
Reconstruction Method:
SINGLE PARTICLE
