Deposition Date
2022-11-22
Release Date
2023-08-16
Last Version Date
2024-11-06
Entry Detail
PDB ID:
8F8N
Keywords:
Title:
Crystal structure of the Arabidopsis SPIRAL2 C-terminal domain
Biological Source:
Source Organism(s):
Arabidopsis thaliana (Taxon ID: 3702)
Expression System(s):
Method Details:
Experimental Method:
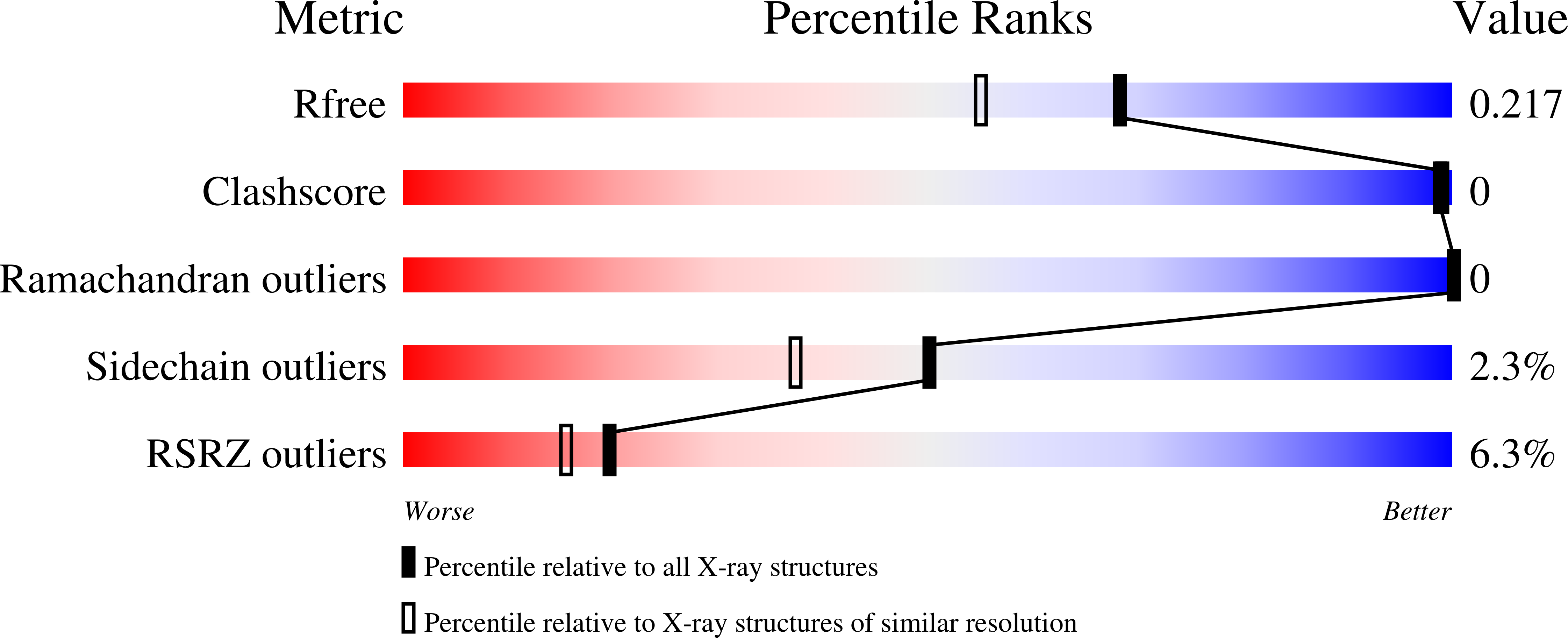
Resolution:
1.80 Å
R-Value Free:
0.21
R-Value Work:
0.18
R-Value Observed:
0.18
Space Group:
P 21 21 21