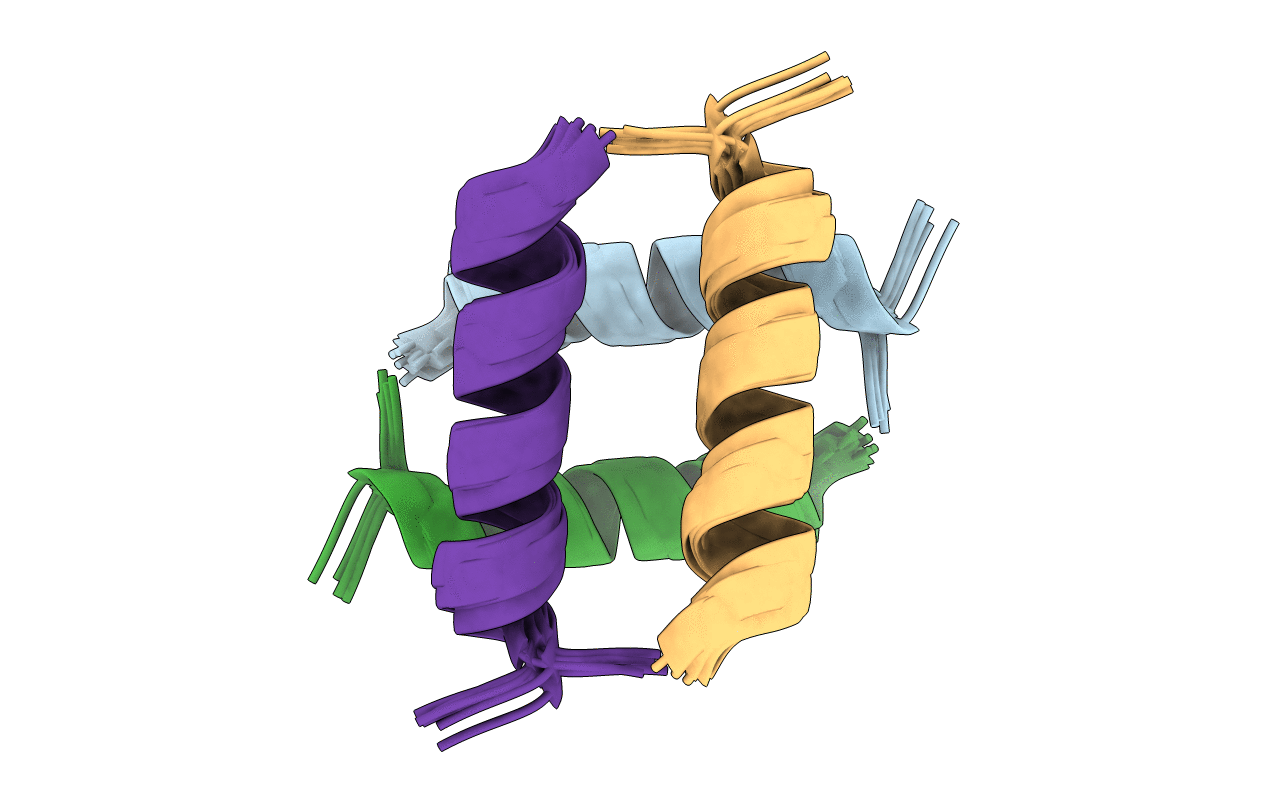
Deposition Date
2018-11-29
Release Date
2019-02-13
Last Version Date
2024-05-01
Entry Detail
PDB ID:
6N8C
Keywords:
Title:
Structure of the Huntingtin tetramer/dimer mixture determined by paramagnetic NMR
Biological Source:
Source Organism(s):
Homo sapiens (Taxon ID: 9606)
Expression System(s):
Method Details:
Experimental Method:
Conformers Calculated:
1100
Conformers Submitted:
10
Selection Criteria:
structures with the lowest energy