Deposition Date
2018-11-30
Release Date
2019-01-09
Last Version Date
2024-11-20
Entry Detail
PDB ID:
6IBL
Keywords:
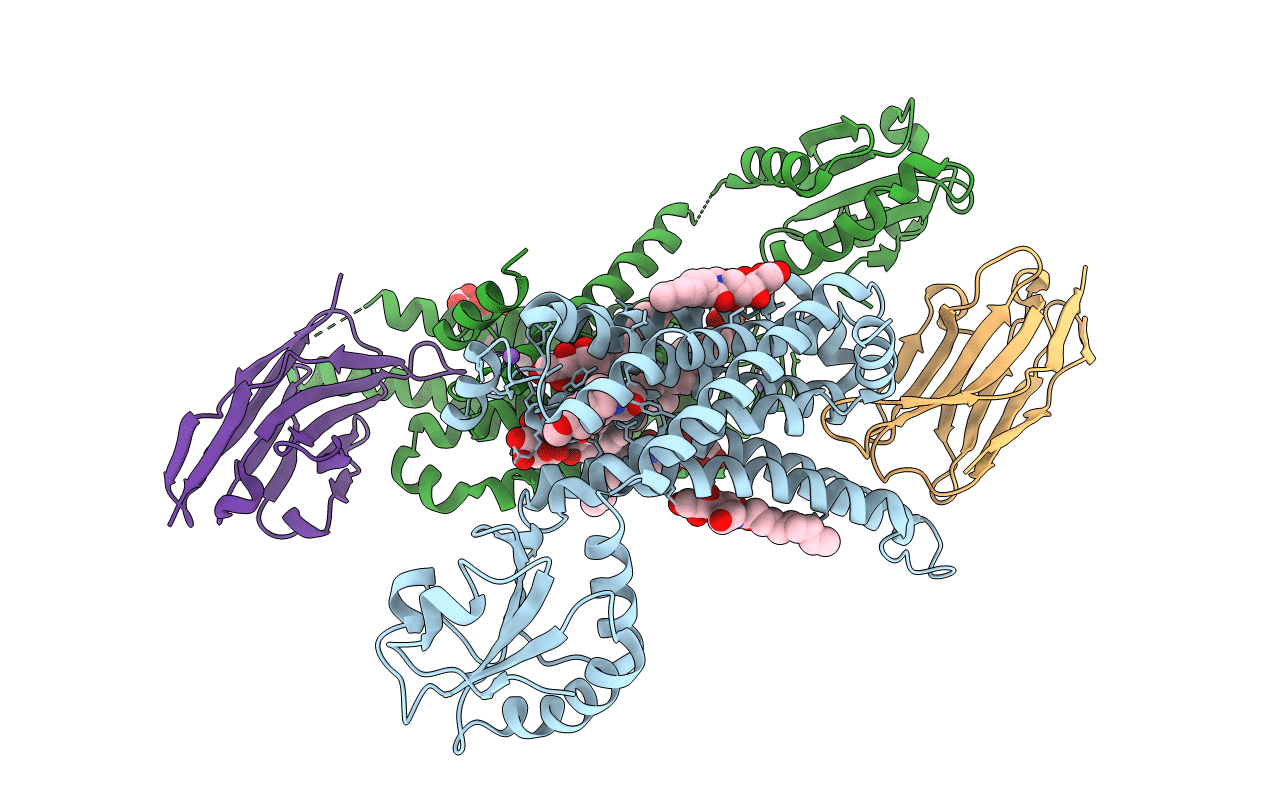
Title:
ACTIVATED TURKEY BETA1 ADRENOCEPTOR WITH BOUND AGONIST FORMOTEROL AND NANOBODY Nb80
Biological Source:
Source Organism(s):
Escherichia coli (strain K12) (Taxon ID: 83333)
Meleagris gallopavo (Taxon ID: 9103)
Lama glama (Taxon ID: 9844)
Meleagris gallopavo (Taxon ID: 9103)
Lama glama (Taxon ID: 9844)
Expression System(s):
Method Details:
Experimental Method:
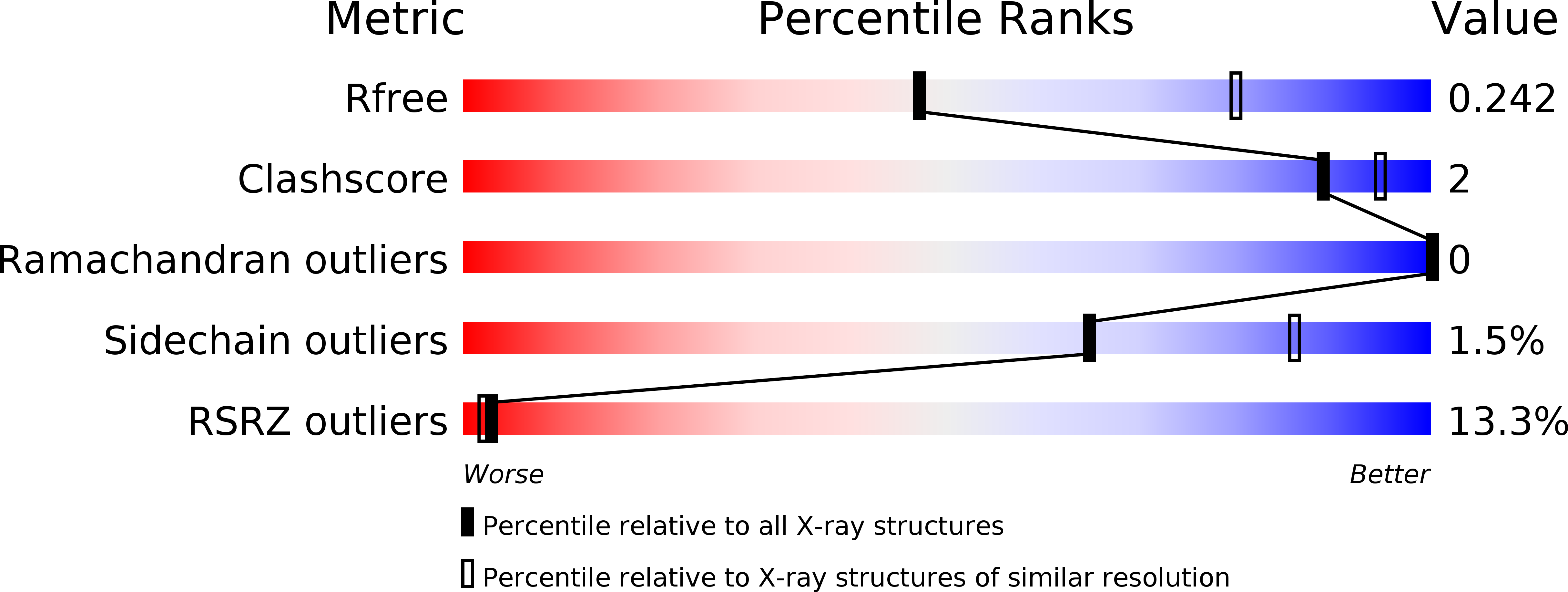
Resolution:
2.70 Å
R-Value Free:
0.27
R-Value Work:
0.23
R-Value Observed:
0.24
Space Group:
P 21 21 21