Deposition Date
2016-07-15
Release Date
2016-09-21
Last Version Date
2023-10-04
Entry Detail
PDB ID:
5KVQ
Keywords:
Title:
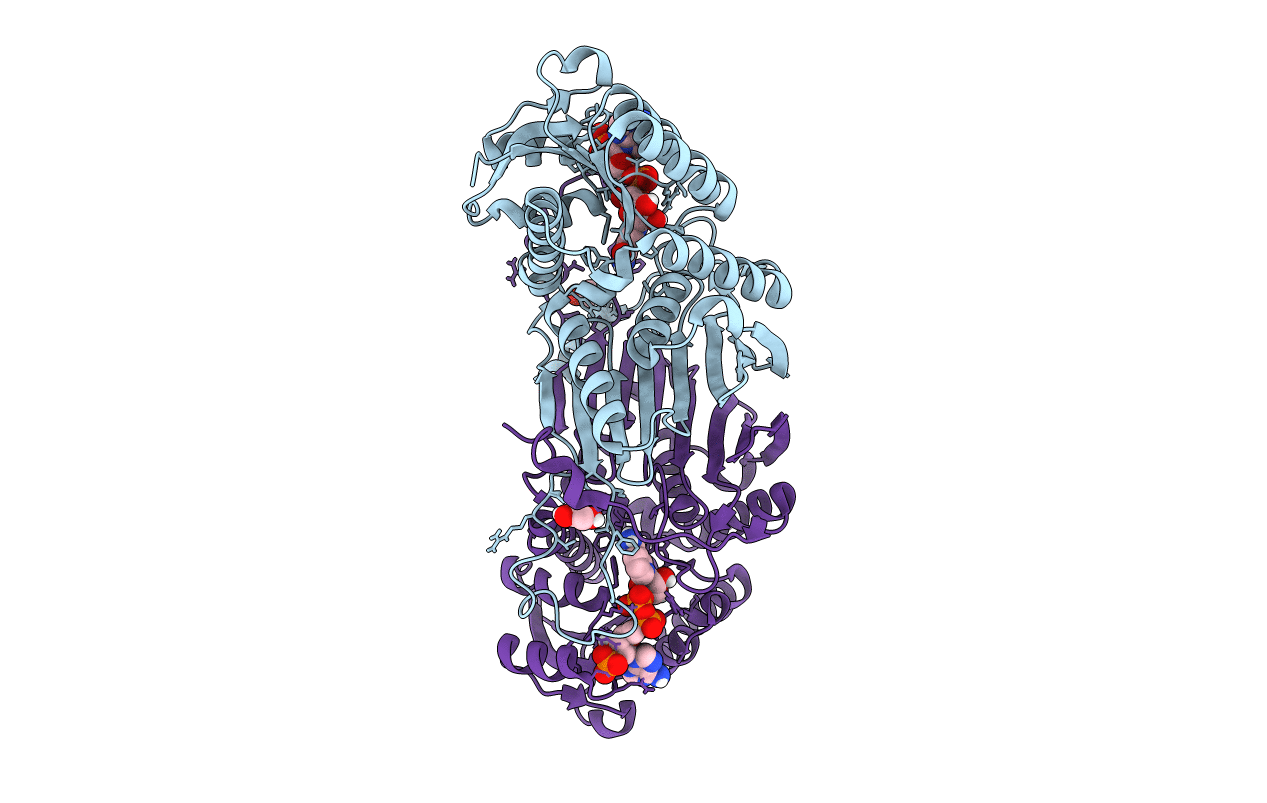
NADP+ bound structure of Irp3, a Thiazolinyl Imine Reductase from Yersinia enterocolitica
Biological Source:
Source Organism(s):
Yersinia enterocolitica (Taxon ID: 630)
Expression System(s):
Method Details:
Experimental Method:
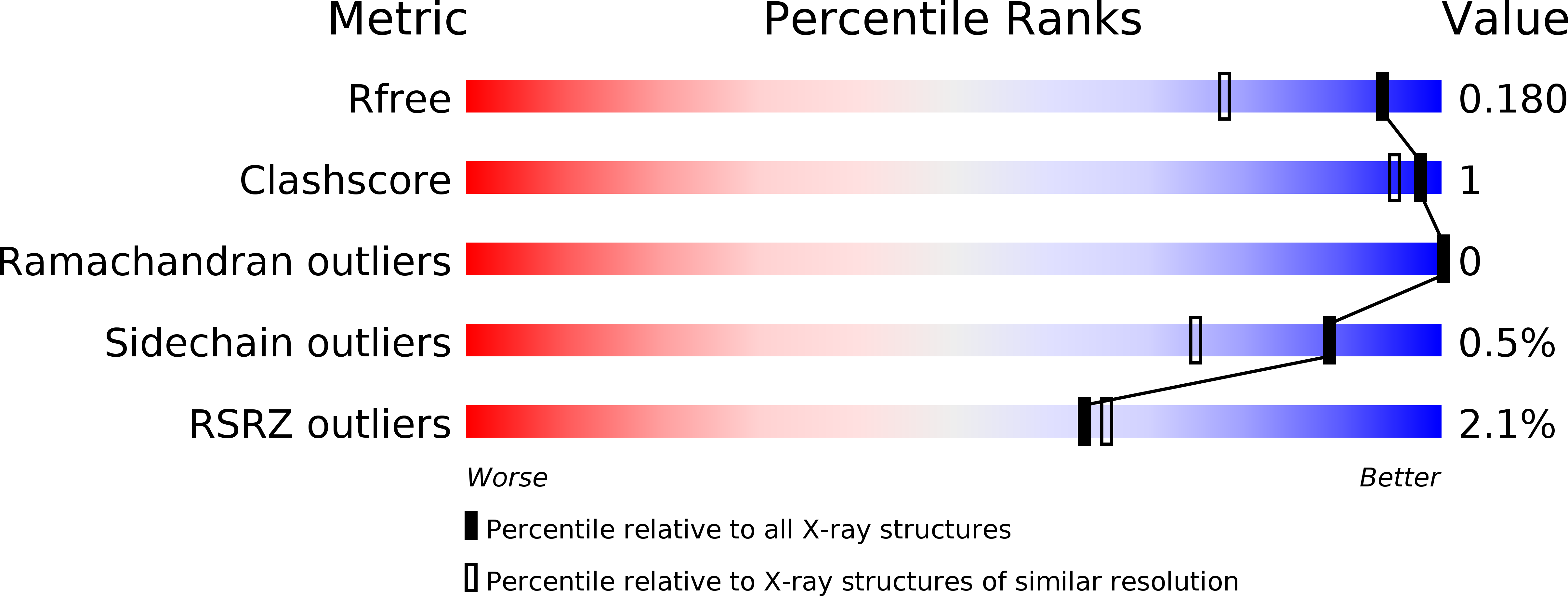
Resolution:
1.45 Å
R-Value Free:
0.17
R-Value Work:
0.15
R-Value Observed:
0.15
Space Group:
P 1 21 1