
Deposition Date
2015-06-19
Release Date
2016-06-29
Last Version Date
2024-10-16
Entry Detail
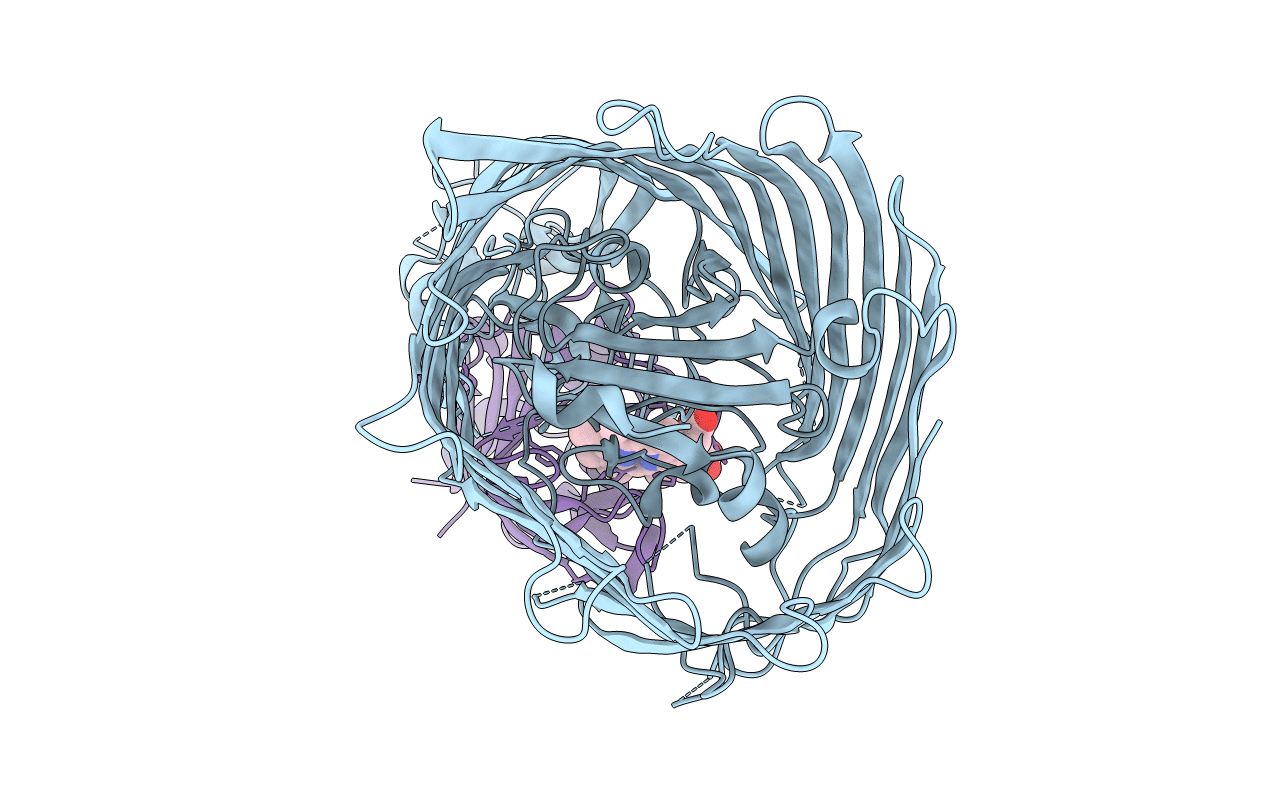
PDB ID:
5C58
Keywords:
Title:
A double mutant of serratia marcescens hemophore receptor HasR in complex with its hemophore HasA and heme
Biological Source:
Source Organism(s):
Serratia marcescens (Taxon ID: 615)
Expression System(s):
Method Details:
Experimental Method:
Resolution:
2.80 Å
R-Value Free:
0.29
R-Value Work:
0.24
R-Value Observed:
0.24
Space Group:
I 2 2 2
