Deposition Date
2010-03-25
Release Date
2011-04-06
Last Version Date
2023-09-06
Entry Detail
PDB ID:
3MBC
Keywords:
Title:
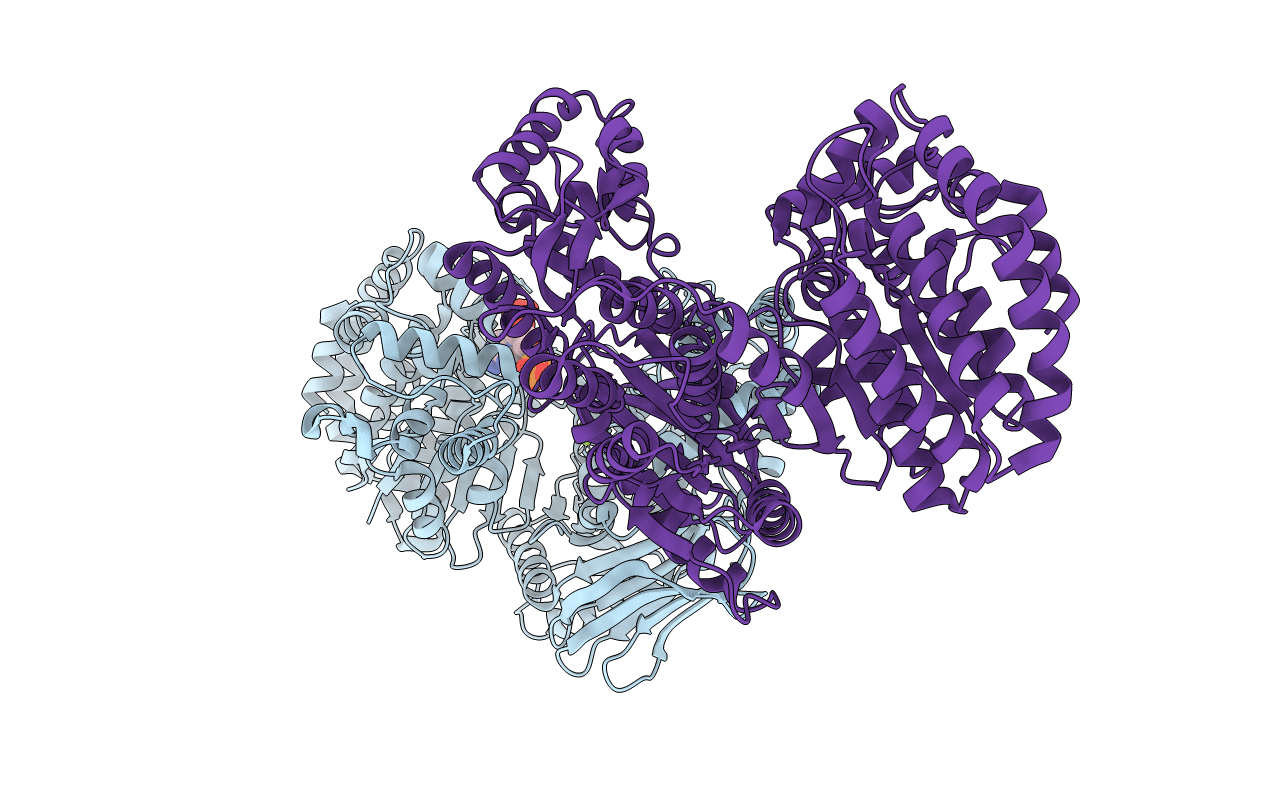
Crystal structure of monomeric isocitrate dehydrogenase from Corynebacterium glutamicum in complex with NADP
Biological Source:
Source Organism(s):
Corynebacterium glutamicum (Taxon ID: 196627)
Method Details:
Experimental Method:
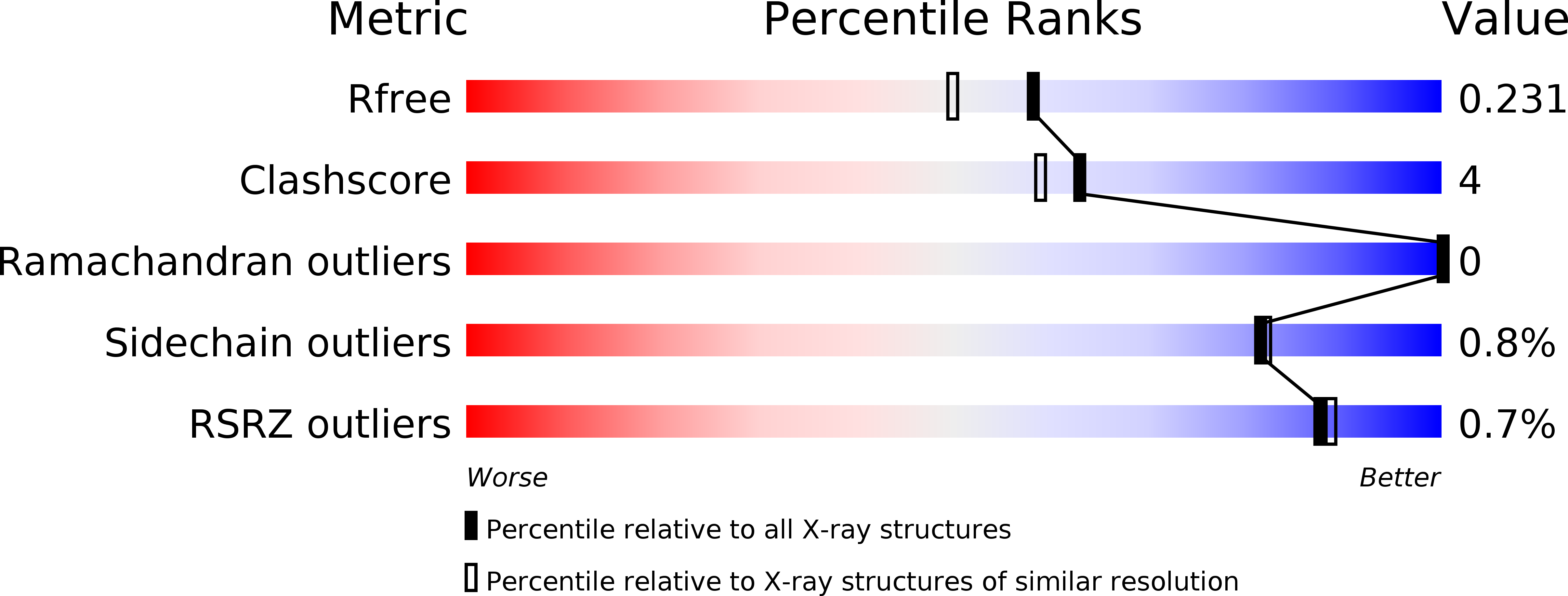
Resolution:
1.90 Å
R-Value Free:
0.22
R-Value Work:
0.18
R-Value Observed:
0.19
Space Group:
C 1 2 1