
Deposition Date
2009-02-02
Release Date
2009-05-05
Last Version Date
2024-10-30
Entry Detail
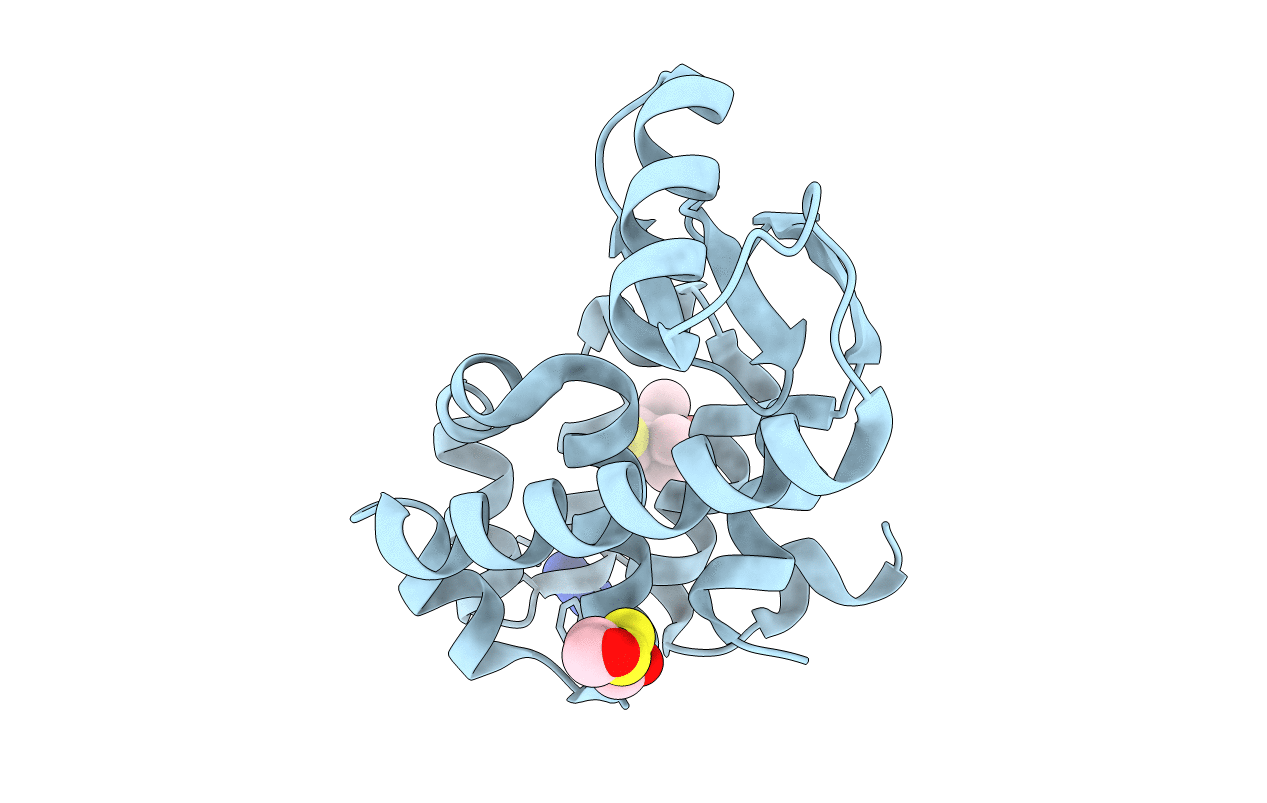
PDB ID:
3G3V
Keywords:
Title:
Crystal structure of spin labeled T4 Lysozyme (V131R1) at 291 K
Biological Source:
Source Organism(s):
Enterobacteria phage T4 (Taxon ID: 10665)
Expression System(s):
Method Details:
Experimental Method:
Resolution:
2.10 Å
R-Value Free:
0.19
R-Value Work:
0.14
R-Value Observed:
0.14
Space Group:
P 32 2 1
