Deposition Date
2010-08-09
Release Date
2010-08-18
Last Version Date
2024-11-20
Entry Detail
PDB ID:
2XO4
Keywords:
Title:
RIBONUCLEOTIDE REDUCTASE Y730NH2Y MODIFIED R1 SUBUNIT OF E. COLI
Biological Source:
Source Organism(s):
ESCHERICHIA COLI (Taxon ID: 83333)
ESCHERICHIA COLI (Taxon ID: 562)
ESCHERICHIA COLI (Taxon ID: 562)
Expression System(s):
Method Details:
Experimental Method:
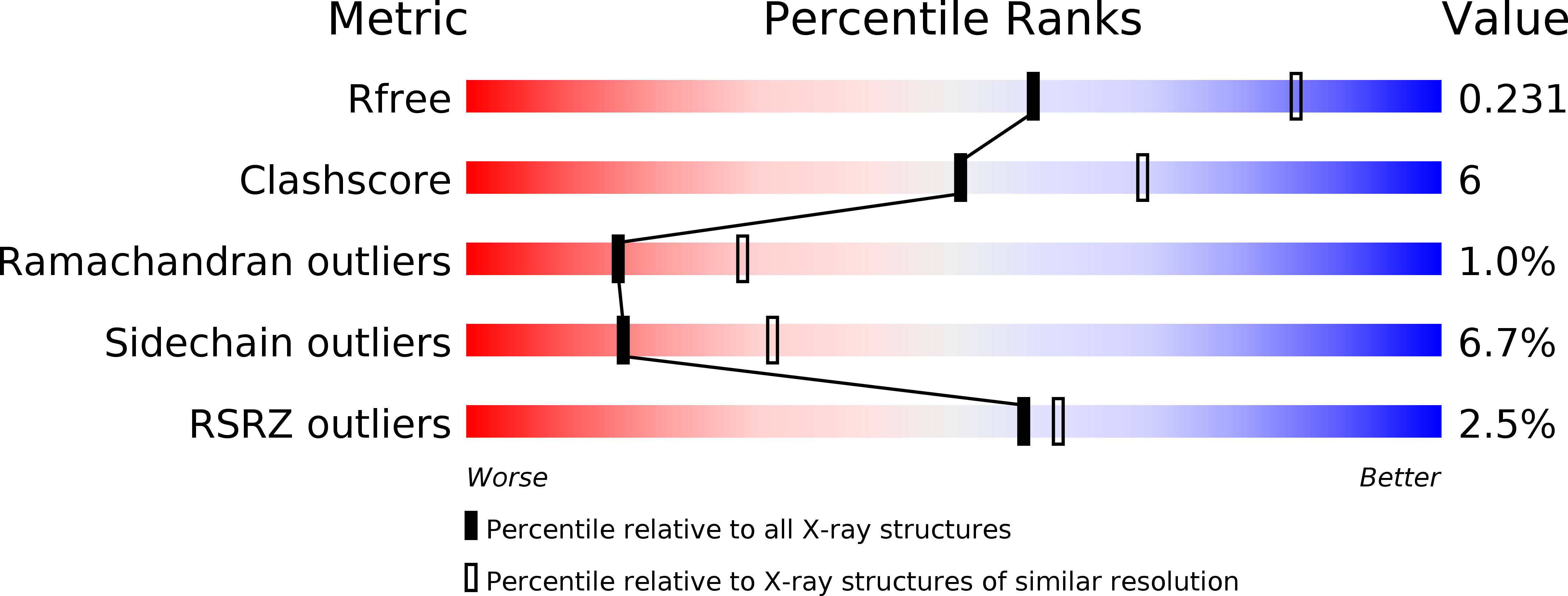
Resolution:
2.50 Å
R-Value Free:
0.23
R-Value Work:
0.18
R-Value Observed:
0.19
Space Group:
H 3 2