Deposition Date
2004-09-17
Release Date
2005-09-27
Last Version Date
2024-11-20
Entry Detail
PDB ID:
1XGY
Keywords:
Title:
Crystal Structure of Anti-Meta I Rhodopsin Fab Fragment K42-41L
Biological Source:
Source Organism(s):
Mus musculus (Taxon ID: 10090)
Method Details:
Experimental Method:
Resolution:
2.71 Å
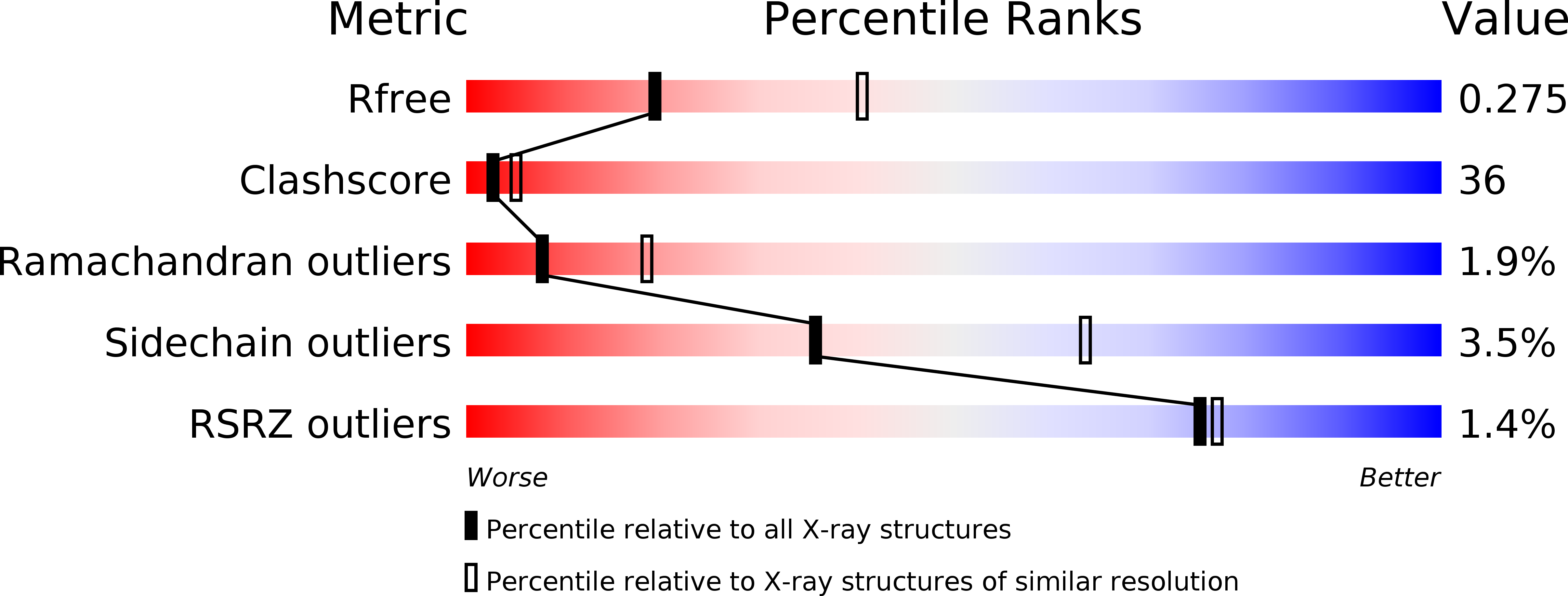
R-Value Free:
0.27
R-Value Work:
0.23
R-Value Observed:
0.23
Space Group:
P 21 21 2