Deposition Date
2003-10-08
Release Date
2004-02-03
Last Version Date
2024-02-14
Entry Detail
PDB ID:
1R4W
Keywords:
Title:
Crystal structure of Mitochondrial class kappa glutathione transferase
Biological Source:
Source Organism(s):
Rattus norvegicus (Taxon ID: 10116)
Expression System(s):
Method Details:
Experimental Method:
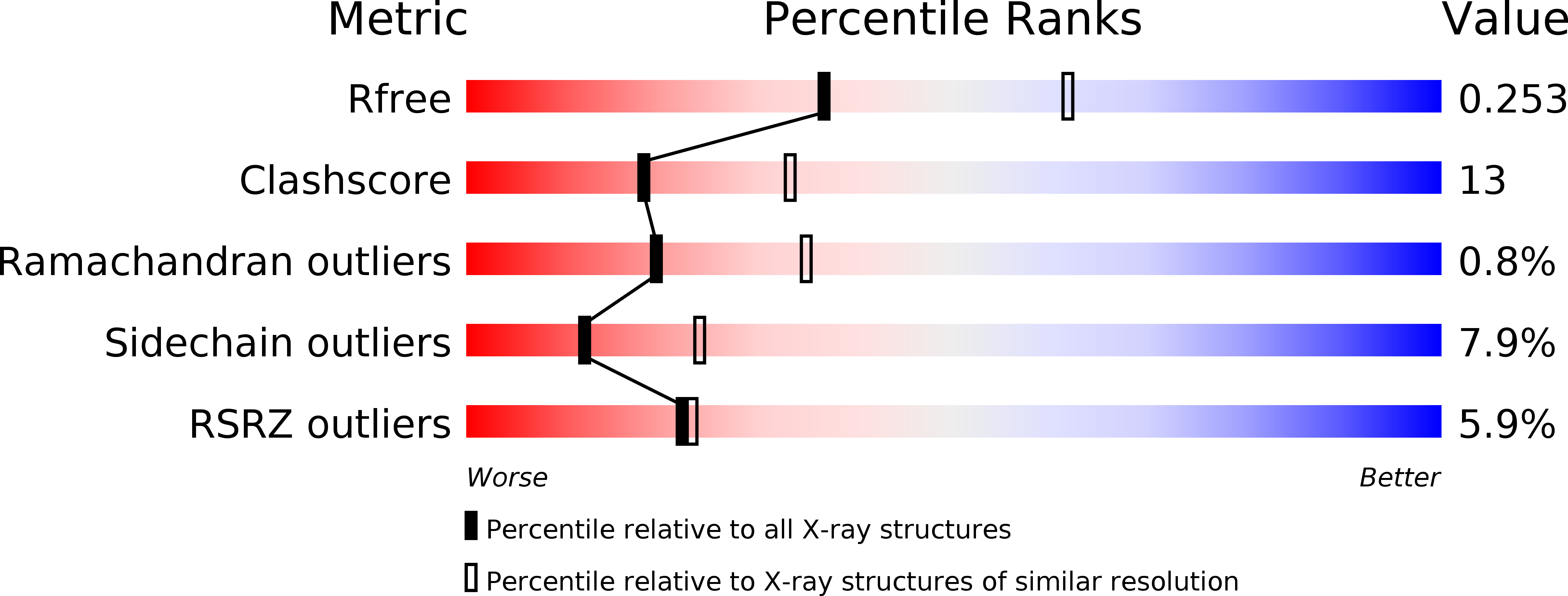
Resolution:
2.50 Å
R-Value Free:
0.25
R-Value Work:
0.20
Space Group:
P 1 21 1