Deposition Date
2002-09-13
Release Date
2002-11-14
Last Version Date
2024-10-23
Entry Detail
Biological Source:
Source Organism:
HUMAN HERPESVIRUS 4 (Taxon ID: 10376)
Host Organism:
Method Details:
Experimental Method:
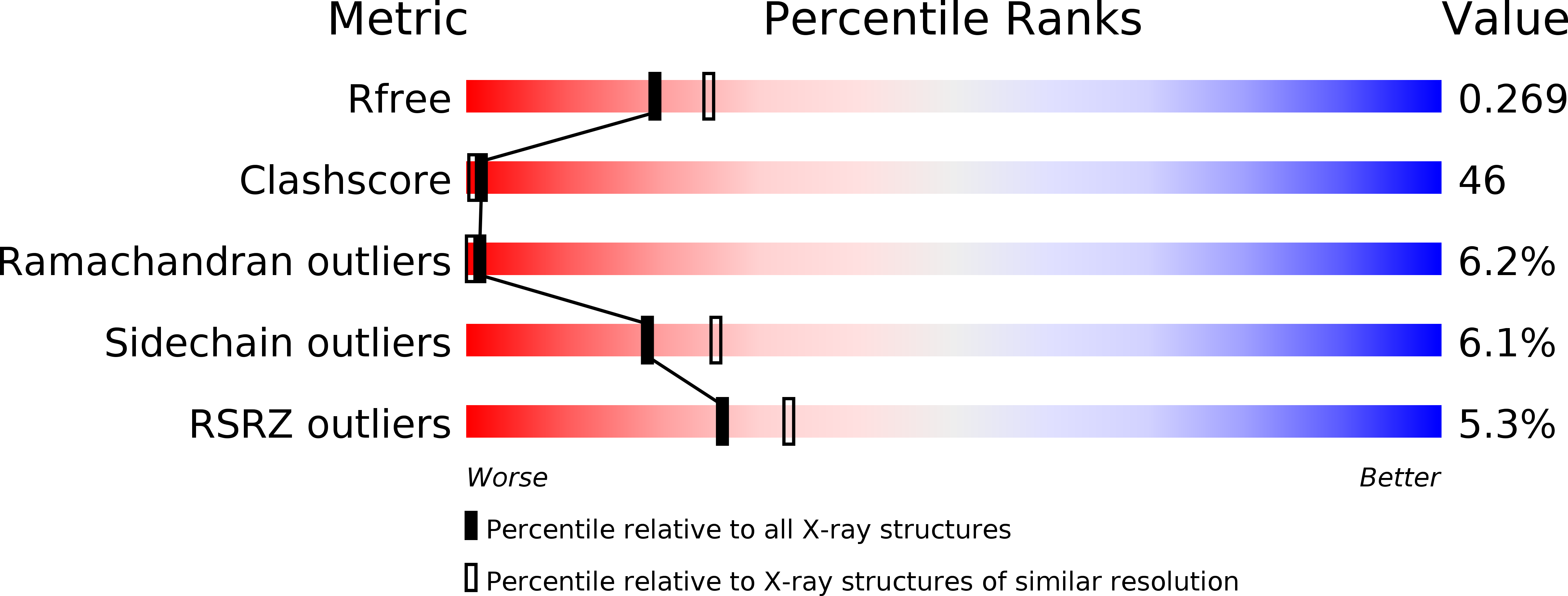
Resolution:
2.30 Å
R-Value Free:
0.27
R-Value Work:
0.19
R-Value Observed:
0.19
Space Group:
P 31 2 1