
Deposition Date
2001-06-19
Release Date
2002-07-05
Last Version Date
2023-08-16
Entry Detail
PDB ID:
1JEZ
Keywords:
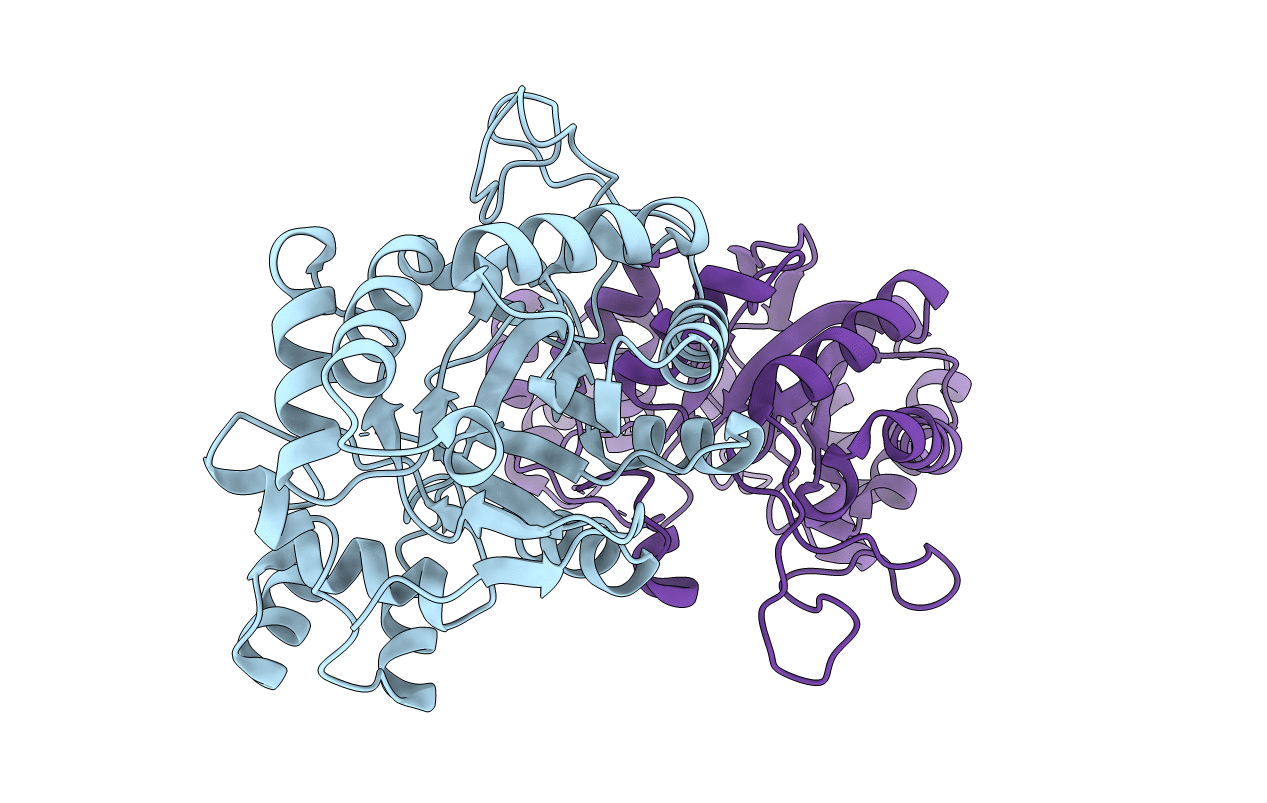
Title:
THE STRUCTURE OF XYLOSE REDUCTASE, A DIMERIC ALDO-KETO REDUCTASE FROM CANDIDA TENUIS
Biological Source:
Source Organism(s):
Candida tenuis (Taxon ID: 45596)
Expression System(s):
Method Details:
Experimental Method:
Resolution:
2.20 Å
R-Value Free:
0.23
R-Value Work:
0.17
R-Value Observed:
0.17
Space Group:
C 1 2 1
