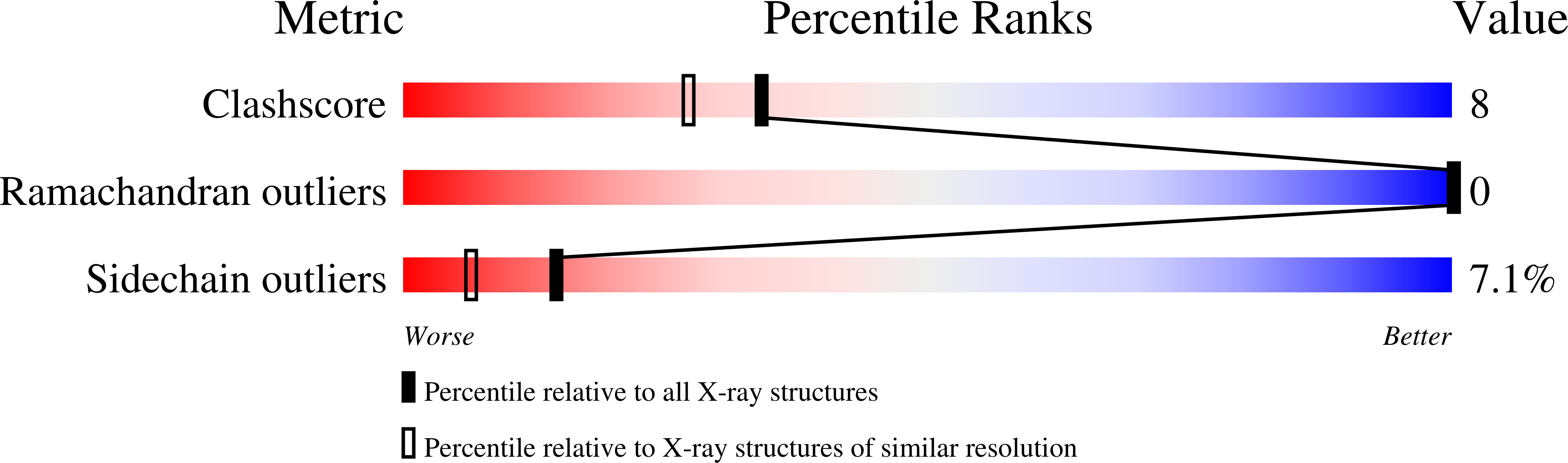
Deposition Date
1997-07-24
Release Date
1998-01-28
Last Version Date
2024-05-22
Entry Detail
PDB ID:
1AP6
Keywords:
Title:
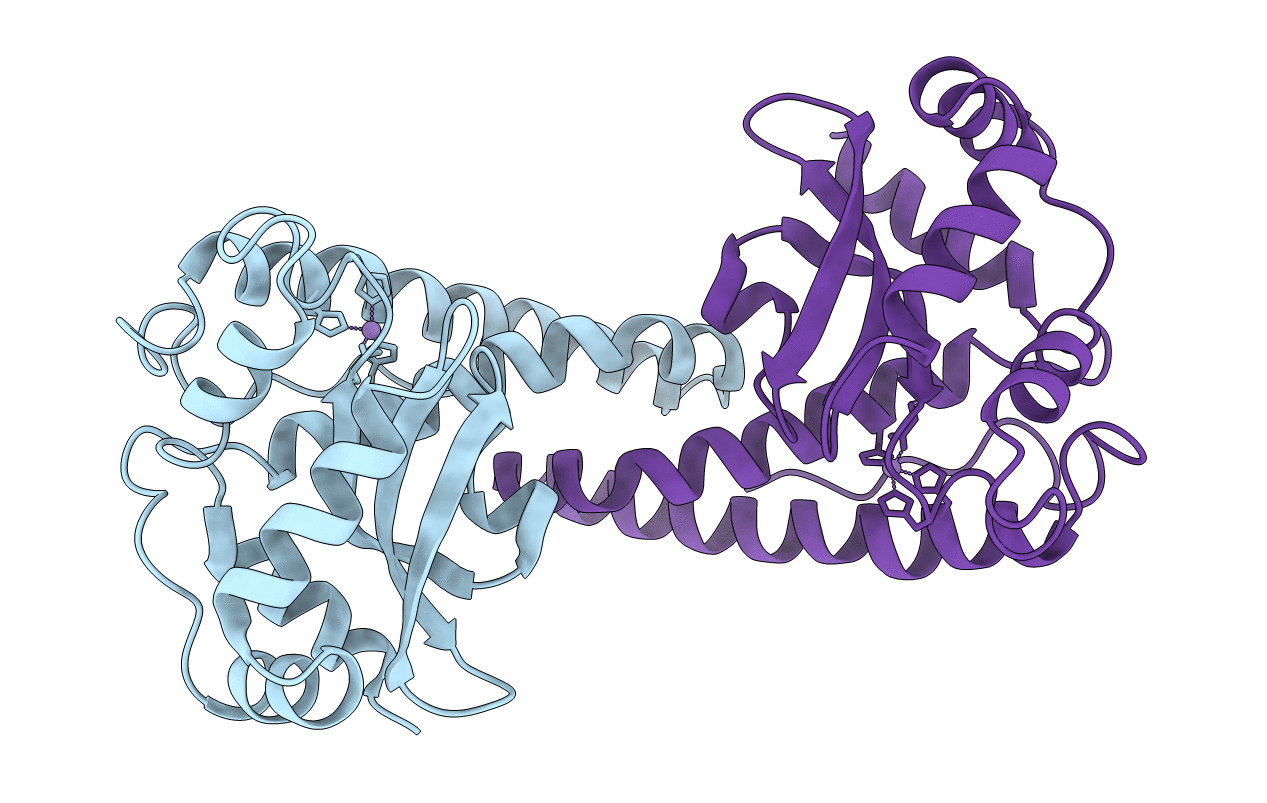
TYR34->PHE MUTANT OF HUMAN MITOCHONDRIAL MANGANESE SUPEROXIDE DISMUTASE
Biological Source:
Source Organism(s):
Homo sapiens (Taxon ID: 9606)
Expression System(s):
Method Details:
Experimental Method:
Resolution:
1.90 Å
R-Value Free:
0.23
R-Value Work:
0.18
R-Value Observed:
0.18
Space Group:
P 61 2 2